京の伝統野菜ミブナの育種の歴史を解明!
「Horticulture Research」オンライン版に掲載
2021.06.02
京都産業大学生態進化発生学研究室の木村成介教授、坂本智昭助教、川勝弥一研究員(研究当時)らの共同研究グループはミブナの誕生に、同じアブラナ科の植物であるカブ類が関わっていた可能性があることを見出し、文献調査と遺伝子解析を組み合わせる研究を進め、伝統野菜の新たな品種であるミブナがミズナから分岐して誕生した過程とその遺伝的背景を明らかにしました。
今後、国内の文献記録に注目することで、日本特有の品種や作物が誕生した過程やその遺伝的背景について、詳細に理解できることが期待されています。
リリース日:2021-06-02
発表論文
「Combination of genetic analysis and ancient literature survey reveals the divergence of traditional Brassica rapa varieties from Kyoto, Japan」
(京の伝統野菜「壬生菜」が「水菜」から分岐して誕生した過程を、遺伝的解析と古文書解析の組み合わせにより解明)
(京の伝統野菜「壬生菜」が「水菜」から分岐して誕生した過程を、遺伝的解析と古文書解析の組み合わせにより解明)
本件のポイント
- 【遺伝的解析】ミズナとミブナの交配実験、および次世代シークエンス技術を用いた遺伝的解析から、ミズナとミブナの葉の形の違いには、BrTCP15という遺伝子が関わっていることが示されました。
- 【古文書解析】日本国内の古文書の記録から、ミブナの誕生にはカブ類との交雑が関わっていた可能性を見出しました。
- そこで、カブ類についても次世代シークエンス解析を行った結果、紫姫という品種のカブもミブナと同じBrTCP15を有していることが明らかになりました。
- これらの結果から、およそ200年前にミズナとカブが交雑したことが、ヘラ型の葉を持つミブナが誕生したきっかけであるとの結論に達しました。
概要
ミブナは京都の壬生地域発祥の京野菜です。ミブナは同じ京野菜であるミズナと同一の種ですが、ミズナの葉は切れ込みのあるギザギザな形、ミブナの葉は単純な丸いヘラ型をしています。
ミブナがミズナの栽培過程で誕生したことは知られていましたが、1787年の古文書『拾遺都名所図会(しゅういみやこめいしょずえ)』に描かれた‘壬生菜’とされる作物には、現在のミズナのような切れ込みのある葉が描かれています。この‘壬生菜’のギザギザな形が、250年足らずの間に、いつ、どのようにして現在のミブナに見られるヘラ型になったのかは不明でした。
京都産業大学の川勝弥一博士研究員(研究当時)、坂本智昭助教、木村成介教授(生態進化発生学研究室)らの共同研究グループは、ミブナの誕生に、同じアブラナ科の植物であるカブ類が関わっていた可能性があることを見出し、これを手掛かりに、今回、他の古文書も参照しながら、文献調査と遺伝子解析を組み合わせる研究を進め、伝統野菜の新たな品種であるミブナがミズナから分岐して誕生した過程とその遺伝的背景を明らかにしました。
ミブナがミズナの栽培過程で誕生したことは知られていましたが、1787年の古文書『拾遺都名所図会(しゅういみやこめいしょずえ)』に描かれた‘壬生菜’とされる作物には、現在のミズナのような切れ込みのある葉が描かれています。この‘壬生菜’のギザギザな形が、250年足らずの間に、いつ、どのようにして現在のミブナに見られるヘラ型になったのかは不明でした。
京都産業大学の川勝弥一博士研究員(研究当時)、坂本智昭助教、木村成介教授(生態進化発生学研究室)らの共同研究グループは、ミブナの誕生に、同じアブラナ科の植物であるカブ類が関わっていた可能性があることを見出し、これを手掛かりに、今回、他の古文書も参照しながら、文献調査と遺伝子解析を組み合わせる研究を進め、伝統野菜の新たな品種であるミブナがミズナから分岐して誕生した過程とその遺伝的背景を明らかにしました。
背景
京都には京野菜と呼ばれる伝統野菜があります。京野菜は、京都が日本の都であった794年から1868年までの間に、天皇への献上品として多くの品種の野菜が各地から集められ、栽培されたことに由来すると考えられています。近年、伝統野菜は地元野菜のブランド化や作物の多様性などの側面から価値が見直されています。今後、伝統野菜の価値をさらに高めるためには、これらの野菜類を科学的に解析し、それらの来歴を明らかにすることが重要と考えられます。
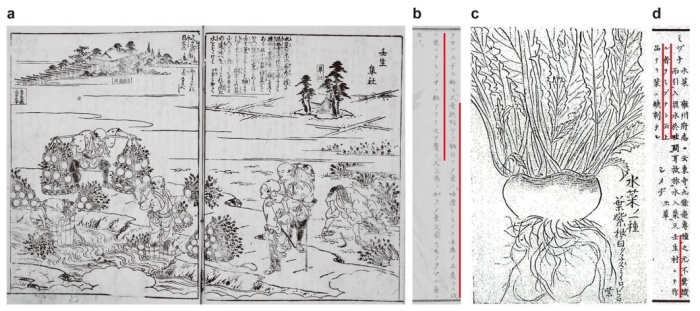
ミズナとミブナは、いずれもよく知られた京野菜です。ミズナは特徴的な鋸歯(きょし)のある葉(ギザギザした形の葉)を有していますが、ミブナは単純なヘラ状の葉を有しています(図1a-d)。両者は葉の形態が全く異なっていますが、分類学的には同一種です。ミブナは京都の壬生地方で、ミズナの栽培過程で生じたと伝えられています。一方、1787年の文献である『拾遺都名所図会』には、現在のミズナのような切れ込みのある葉をもっているミブナが描かれており(図1e)、ミブナの葉の形態が、いつ、どのようにして現在のヘラ型になったのかは明らかにされていませんでした。
ミズナとミブナは、いずれもよく知られた京野菜です。ミズナは特徴的な鋸歯(きょし)のある葉(ギザギザした形の葉)を有していますが、ミブナは単純なヘラ状の葉を有しています(図1a-d)。両者は葉の形態が全く異なっていますが、分類学的には同一種です。ミブナは京都の壬生地方で、ミズナの栽培過程で生じたと伝えられています。一方、1787年の文献である『拾遺都名所図会』には、現在のミズナのような切れ込みのある葉をもっているミブナが描かれており(図1e)、ミブナの葉の形態が、いつ、どのようにして現在のヘラ型になったのかは明らかにされていませんでした。
研究成果
手法1:遺伝的解析
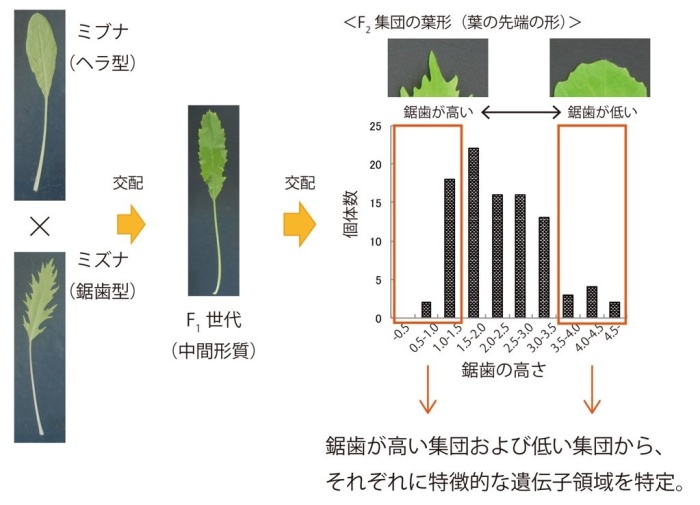
この課題に関心をもった私たちは、ミズナからミブナが誕生した背景を解明するために、最も大きな形態的な違いである「葉の形」について遺伝的解析を行いました。まず、ミズナとミブナを交配させ、得られたF1世代*1を自家受粉させることによってF2世代*2を得ました(図2)。このF2世代の葉は、ミズナのように鋸歯が大きなものから、ミブナのようなヘラ型に近いものまでさまざまな形が見られました。
次に、ゲノムシークエンス解析*3技術を用いた遺伝子解析を行いました。F2世代の葉の鋸歯が高い集団と低い集団について、それぞれの集団に特徴的な遺伝子領域を特定するQTL-seqという解析を行いました(図2)。その結果、ミブナには7番染色体*4の一部の配列がミズナとは異なっており、その領域からはTEOSINTE BRANCHED/CYCLOIDEA/PCF15 (BrTCP15) という遺伝子が見つかりました。
BrTCP15遺伝子は、葉の形態形成に寄与することがシロイヌナズナ*5の研究から知られています。よって私たちは、「ミズナとミブナの葉の形態の違いの主な原因はBrTCP15 の配列の違いによる」と結論づけました。
次に、ゲノムシークエンス解析*3技術を用いた遺伝子解析を行いました。F2世代の葉の鋸歯が高い集団と低い集団について、それぞれの集団に特徴的な遺伝子領域を特定するQTL-seqという解析を行いました(図2)。その結果、ミブナには7番染色体*4の一部の配列がミズナとは異なっており、その領域からはTEOSINTE BRANCHED/CYCLOIDEA/PCF15 (BrTCP15) という遺伝子が見つかりました。
BrTCP15遺伝子は、葉の形態形成に寄与することがシロイヌナズナ*5の研究から知られています。よって私たちは、「ミズナとミブナの葉の形態の違いの主な原因はBrTCP15 の配列の違いによる」と結論づけました。
手法2:古文書調査
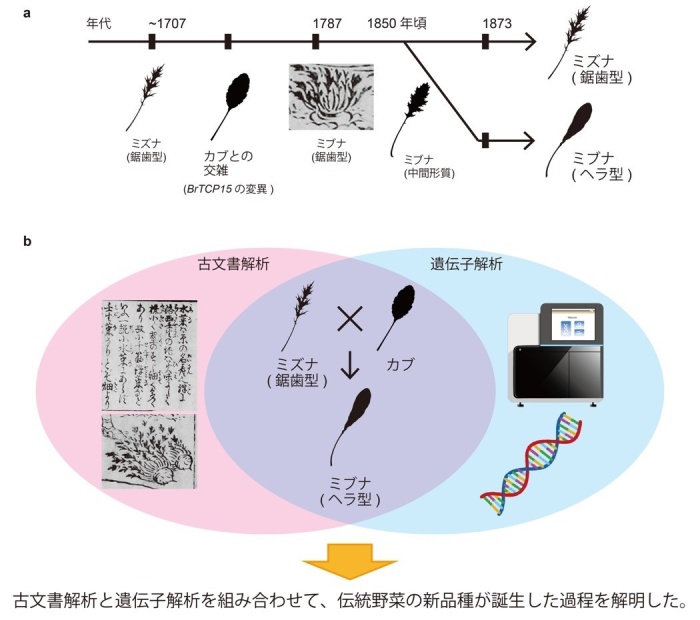
次に、ミズナからミブナが分岐した育種の過程を明らかにするために、古文書の調査を行いました。最も古いミブナの記録は1787年に刊行された『拾遺都名所図会』で見つかり、ここで描かれているミブナは、現在のミズナのような切れ込みのある葉が描かれていました(図3a)。以後のミブナの記録では、1850年頃に刊行された『草木図説』には「ミブナの葉は欠刻(=ギザギザ)が少ない」と記されていた他(図3b)、ほぼ同時期の文献である『植物図説雑纂』には、ミズナの一種としてやや単純な葉を持つ植物の記録が残されていました(図3c)。さらに、1873年に刊行された『日本産物誌』には「ミブナの葉に欠刻なし」と記されていました(図3d)。以上より、「ミブナは1800年代中頃から葉の形態が単純化し始めて、1800年代後半にはヘラ型の葉として定着した」と考えられました。
ここで、ミズナからミブナへの移行期間に刊行された『植物図説雑纂』のミズナの一種に再度注目すると、この野菜はカブのような大きな胚軸*6を持つことがわかります(図3c)。さらに、最古のミブナの記録である上述の『拾遺都名所図会』に描かれたミブナも、カブのような胚軸が描かれていました(図1e)。そこで、「ミズナとカブとが交雑したことが、現在のヘラ型の葉を持つミブナが誕生したきっかけである」という仮説を立てました。
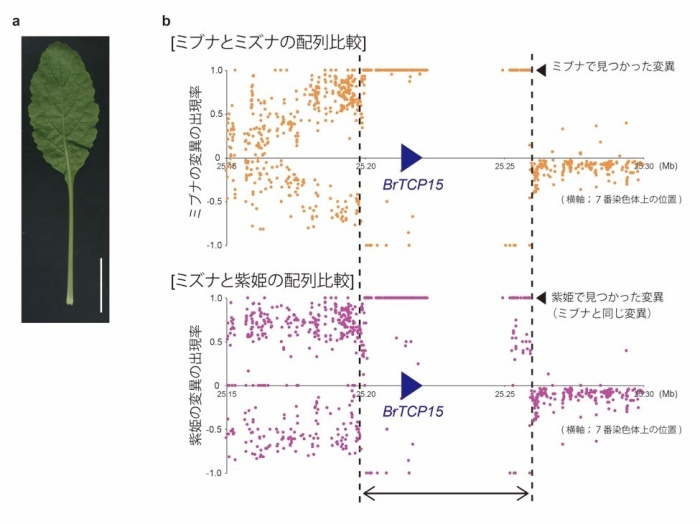
この仮説を検証するために、カブ類についてもゲノム配列の解析を行い、ミズナとミブナの配列との比較を行いました。その結果、紫姫という品種のカブのゲノムは、ミズナとミブナの葉の形の違いに関わるBrTCP15遺伝子を含む領域が、ミブナに近い配列であることがわかりました(図4a-b)。この結果は、ミブナのヘラ型の葉の形態形成に関わる遺伝子が、カブ類に由来していた可能性を示しています。
ここで、ミズナからミブナへの移行期間に刊行された『植物図説雑纂』のミズナの一種に再度注目すると、この野菜はカブのような大きな胚軸*6を持つことがわかります(図3c)。さらに、最古のミブナの記録である上述の『拾遺都名所図会』に描かれたミブナも、カブのような胚軸が描かれていました(図1e)。そこで、「ミズナとカブとが交雑したことが、現在のヘラ型の葉を持つミブナが誕生したきっかけである」という仮説を立てました。
この仮説を検証するために、カブ類についてもゲノム配列の解析を行い、ミズナとミブナの配列との比較を行いました。その結果、紫姫という品種のカブのゲノムは、ミズナとミブナの葉の形の違いに関わるBrTCP15遺伝子を含む領域が、ミブナに近い配列であることがわかりました(図4a-b)。この結果は、ミブナのヘラ型の葉の形態形成に関わる遺伝子が、カブ類に由来していた可能性を示しています。
今後の展望
以上の結果から、現在のヘラ型の葉のミブナの誕生の原因は、1800年頃にミズナとカブとが交雑したことである可能性が示されました(図5a-b)。葉の形態の多様性は、光合成効率を高めるためや、環境適応のためなどの理由で生じたと考えられていますが、一方で、栽培の効率化や食事をより豊かにするために、人為的に多様化が進められたことも知られています。国内の文献記録に注目することで、日本特有の品種や作物が誕生した過程やその遺伝的背景について、詳細に理解できることが期待できます。
責任著者コメント
今回、京都の伝統野菜であるミズナとミブナの葉の形態の研究により、ミブナは、ミズナとカブとが交雑したことによって誕生したことが示されました。京野菜については、国内に先人たちの手による多数の記録が残されており、これらの調査から推察された育種の過程を、生命科学的な手法(遺伝的解析)によって裏付けることができました。
古文書が残っているのは京都だけではありません。日本国内には、野菜、花、動物など、さまざまな生物の記録が多数残されています。これらの調査と生命科学的な手法を組み合わせることで、今後、日本特有の品種や作物について詳細な理解が進んでゆくと考えています。今回の研究成果を通して、野菜類をはじめとした生物の多様性や形態の特徴などに関心を持つきっかけにしていただければと願っています。
(京都産業大学 木村成介)
古文書が残っているのは京都だけではありません。日本国内には、野菜、花、動物など、さまざまな生物の記録が多数残されています。これらの調査と生命科学的な手法を組み合わせることで、今後、日本特有の品種や作物について詳細な理解が進んでゆくと考えています。今回の研究成果を通して、野菜類をはじめとした生物の多様性や形態の特徴などに関心を持つきっかけにしていただければと願っています。
(京都産業大学 木村成介)
用語・事項の解説
1. F1世代
雑種第一代のこと。異なる両親の交雑の結果生じた、第一世代目の子孫。
2. F2世代
雑種第二代のこと。F1世代を自家受粉することで得られる世代。
3. ゲノムシークエンス解析
一つの生物が持つすべての遺伝情報(ゲノム)を、次世代シーケンサーとスーパ-コンピュータを用いることで、網羅的に解析する手法。
4. 染色体
遺伝情報がつまったDNAが太く折り畳まれたもの。ミズナ・ミブナには10対の染色体が存在する。
5. シロイヌナズナ
植物のモデル生物。植物の生命現象の研究に広く用いられている。
6. 胚軸
植物の根と茎の間に位置する器官。カブの可食部の大部分はこの組織に由来する。
参考図

論文情報
| 論文タイトル | Combination of genetic analysis and ancient literature survey reveals the divergence of traditional Brassica rapa varieties from Kyoto, Japan (京の伝統野菜「壬生菜」が「水菜」から分岐して誕生した過程を、遺伝的解析と古文書解析の組み合わせにより解明) |
|---|---|
| 掲載誌 | 国際学術誌「Horticulture Research」オンライン版 |
| 掲載日 | 2021年6月2日(水)(日本時間) |
| 著者 | (1筆頭著者、2責任著者) 1Yaichi Kawakatsu, Tomoaki Sakamoto, Hokuto Nakayama, Kaori Kaminoyama, Kaori Igarashi, Masaki Yasugi, Hiroshi Kudoh, Atsushi J. Nagano, Kentaro Yano, Nakao Kubo, Michitaka Notaguchi, and 2 Seisuke Kimura |
| DOI | 10.1038/s41438-021-00569-0 |
研究者一覧
- 京都産業大学 生命科学部 産業生命科学科
木村成介(教授)、坂本智昭(助教)、上ノ山香織(技術専門職員) - 名古屋大学 生物機能開発利用センター
野田口理孝(准教授)、川勝弥一(博士研究員) - 東京大学 大学院理学系研究科
中山北斗(助教) - 明治大学 農学部
矢野健太郎(教授)、五十嵐香理(博士研究員) - 宇都宮大学 工学部
八杉公基(産学官連携研究員) - 京都大学 生態学研究センター
工藤洋(教授) - 龍谷大学 農学部 植物生命科学科
永野惇(准教授) - 京都府立大学 生命環境科学研究科
久保中央(教授)
謝辞
本研究は、文部科学省および日本学術振興会の科学研究費補助金(16H01472, 18H04787, 19H04870)文部科学省の私立大学戦略的研究基盤形成支援事業(S1511023)、笹川科学研究助成、および明治大学の計算ソフトウェア支援プログラムによる研究費の支援を受けて実施しました。本研究は、情報・システム研究機構 国立遺伝学研究所が有する遺伝研スーパーコンピュータシステムを利用しました。
- お問い合わせ先
-
京都産業大学 広報部
〒603‐8555 京都市北区上賀茂本山
Tel.075-705-1411
kouhou-bu@star.kyoto-su.ac.jp