愮梩巙怣弝嫵庼傜偑丄崙嵺嫟摨尋媶偵傛傝僞儞僷僋幙崌惉憰抲儕儃僜乕儉偲怴惗僞儞僷僋幙偲偺憡屳嶌梡偺徻嵶傪夝柧
丂憤崌惗柦壢妛晹愮梩巙怣弝嫵庼傜偺尋媶僌儖乕僾偑丄僪僀僣丒儈儏儞僿儞戝妛Daniel Wilson攷巑傜偲偺崙嵺嫟摨尋媶偵傛傝丄僞儞僷僋幙崌惉憰抲偱偁傞儕儃僜乕儉偲丄儕儃僜乕儉偵傛偭偰崌惉偝傟偮偮偁傞怴惗僞儞僷僋幙偲偺娫偱婲偙傞憡屳嶌梡傪徻嵶偵夝柧偟傑偟偨丅崱夞偺尋媶惉壥偼丄怴惗僞儞僷僋幙偑帺恎偺崌惉傪惂屼偡傞偨傔偵偳偺傛偆側偐偨偪偱儕儃僜乕儉偵摥偒偐偗偰偄傞偺偐傪柧傜偐偵偟偰偍傝丄嵟嬤條乆側惗暔偱尒弌偝傟偮偮偁傞丄乽僞儞僷僋幙偑帺傜偺惗抋偲塣柦寛掕傪惂屼偡傞乿偲尵偆怴偨側僞儞僷僋幙偺摥偒曽傪傛傝怺偔棟夝偡傞偙偲偵峷專偡傞偲偲傕偵丄堚揱忣曬偐傜惗柦偑抋惗偡傞夁掱偺偆偪丄僞儞僷僋幙偺嵔帺懱偑嶌傜傟偰惗柦婡擻傪妉摼偟偰偄偔巇慻傒傪棟夝偡傞偨傔偺庤偑偐傝偲側傞偙偲偑婜懸偝傟傑偡丅
丂杮尋媶惉壥偼丄2015擭4寧23晅偱丄塸崙嶨帍乽Nature Communications乿偵僆儞儔僀儞宖嵹偝傟傑偟偨丅
宖嵹榑暥柤
丂乽Structure of the Bacillus subtilis 70S ribosome reveals the basis for species-specific stalling乿乮屚憪嬠70S儕儃僜乕儉偺峔憿偐傜尒偊偰偒偨庬摿堎揑東栿掆巭偺暘巕婎斦乯
挊幰
丂Daniel Sohmen乮儈儏儞僿儞戝妛乯丄愮梩巙怣乮嫗嶻戝乯丄愮梩乮壓愳乯捈旤乮嫗嶻戝乯丄C. Axel Innis乮儃儖僪乕戝妛乯丄Otto Berninghausen乮儈儏儞僿儞戝妛乯丄 Roland Beckmann乮儈儏儞僿儞戝妛乯丄埳摗堐徍乮嫗嶻戝乯丄Daniel N. Wilson乮儈儏儞僿儞戝妛乯
尋媶奣梫
攚宨
丂僞儞僷僋幙崌惉憰抲偱偁傞儕儃僜乕儉偼丄儊僢僙儞僕儍乕RNA偵揮幨偝傟偨堚揱忣曬傪愝寁恾偲偟丄偦偺巜椷偵廬偭偰傾儈僲巁摨巑傪寢崌偝偣傞偙偲偱僞儞僷僋幙傪崌惉偟傑偡丅嵶朎撪偱婲偙傞惗壔妛斀墳偼丄懡偔偑僞儞僷僋幙偵傛偭偰怗攠偝傟傞偨傔丄東栿偲屇偽傟傞偙偺僞儞僷僋幙崌惉偺僾儘僙僗偼丄乽惗柦偺愝寁恾偐傜惗柦妶摦偺扴偄庤傪惗傒弌偡夁掱乿偱偁傞偲尵偊傑偡丅捠忢丄僞儞僷僋幙偼丄儕儃僜乕儉偱姰慡偵崌惉偝傟偨屻丄棫懱揑峔憿傊偺愜傝偨偨傒傗摿掕偺応強傊偺堏摦丄懠偺惉暘偲偺夛崌側偳偺夁掱丄偡側傢偪乬惉弉壔乭偺夁掱傪傊偰婡擻傪敪婗偱偒傞忬懺偵側傝傑偡丅偲偙傠偑丄埲慜巹偨偪偑屚憪嬠偱敪尒偟偨MifM偲偄偆僞儞僷僋幙偼丄東栿怢挿偺搑忋偱儕儃僜乕儉偵摥偒偐偗丄帺傜偺崌惉傪堦帪掆巭偝偣傞儐僯乕僋側惈幙傪帩偭偰偄傑偡丅MifM偼丄偙偺東栿偺堦帪掆巭乮東栿傾儗僗僩乯偲偄偆婡擻傪東栿搑忋偱敪婗偡傞丄偡側傢偪東栿搑忋偺忬懺偱摥偔捒偟偄僞儞僷僋幙偱偁傞偙偲偑丄埲慜偺巹偨偪偺尋媶偐傜柧傜偐偵偝傟傑偟偨丅MifM偼丄僞儞僷僋幙偺枌慻崬憰抲偺妶惈傪帺傜偑婎幙偲側偭偰娔帇偟丄憰抲偺摥偒偑掅壓偡傞偲丄戞擇偺僞儞僷僋幙枌慻崬憰抲偺崌惉傪懀恑偡傞偙偲偱丄嵶朎偺妶摦掆巭傪枹慠偵杊偖偲偄偆丄廳梫側栶妱傪扴偭偰偄傑偡丅MifM埲奜偵傕丄東栿傾儗僗僩傪婲偙偡偙偲偱東栿偺搑忋偱摥偔場巕偑條乆側惗暔庬偱尒弌偝傟偰偒偰偍傝丄偙傟傜偺堦楢偺敪尒偼丄僞儞僷僋幙偺婡擻敪尰偺嵼傝曽偵懳偡傞巹偨偪偺棟夝傪奼挘偡傞傕偺偱偟偨丅
丂MifM偺東栿傾儗僗僩偼丄MifM偑崌惉搑忋偵丄帺恎傪崌惉偟偰偄傞儕儃僜乕儉偲摿掕偺憡屳嶌梡傪偡傞偙偲偱婲偙傝傑偡丅埲慜巹偨偪偼丄屚憪嬠偺MifM偼丄戝挵嬠桼棃偺堎庬儕儃僜乕儉偱崌惉偝傟偨偲偒偵偼丄岠棪偺椙偄東栿傾儗僗僩傪堷偒婲偙偡偙偲偑弌棃側偄偙偲傪尒弌偟傑偟偨丅偙偺偙偲偼丄MifM偲儕儃僜乕儉偲偺僐儈儏僯働乕僔儑儞偑丄庬摿堎揑側憡屳嶌梡偵埶懚偡傞傕偺偱偁傞偙偲傪帵偟偰偄傑偡丅偦偙偱丄崱夞丄MifM偵傛傞東栿傾儗僗僩偺儊僇僯僘儉傪棟夝偟丄庬摿堎揑側MifM-儕儃僜乕儉娫憡屳嶌梡偺幚懱傪夝柧偡傞偨傔偵丄儈儏儞僿儞戝妛偺Daniel Wilson攷巑傜偲嫟摨尋媶傪峴偄丄峔憿惗暔妛偲堚揱妛偺庤朄傪梈崌偟偨憤崌揑側傾僾儘乕僠偱丄MifM偺東栿傾儗僗僩偺儊僇僯僘儉偵敆傝傑偟偨丅
尋媶寢壥
丂Daniel Wilson攷巑傜偼丄僋儔僀僆乮掅壏乯揹巕尠旝嬀傪梡偄丄屚憪嬠儕儃僜乕儉偲MifM偺東栿傾儗僗僩忬懺偺暋崌懱偺峔憿傪丄3.5-3.9 Å乮僆儞僌僗僩儘乕儉丄0.1僫僲儊乕僩儖乯偲偄偆崅夝憸搙偱夝柧偟傑偟偨丅偙傟偼丄僌儔儉梲惈嬠仸1 偺儕儃僜乕儉偺峔憿偲偟偰偼丄尰帪揰偱嵟傕崅夝憸搙偺峔憿僨乕僞偱偡丅偙偺暋崌懱偺峔憿夝愅偐傜丄MifM偲儕儃僜乕儉偲偑愙偟偰偄傞偲巚傢傟傞晹暘偑婔偮傕柧傜偐偵側傝傑偟偨乮恾丂嶲徠乯丅儕儃僜乕儉偵偼丄崌惉偝傟偨怴惗僞儞僷僋幙乮怴惗億儕儁僾僠僪嵔乯傪攔弌偡傞僩儞僱儖偑偁傝傑偡丅崱夞丄偙偺僩儞僱儖偺撪暻偵業弌偟偰偄傞L4偲L22偲屇偽傟傞僞儞僷僋幙惉暘偐傜側傞嫹嶓晹埵乮僩儞僱儖偺嫹偔側偭偨晹埵乯偑丄東栿傾儗僗僩偵廳梫側栶妱傪傕偮偙偲偑丄嫗嶻戝僠乕儉偺堚揱妛揑夝愅偐傜晜偐傃忋偑傝傑偟偨丅堦曽丄儈儏儞僿儞戝僠乕儉偵傛傞峔憿夝愅偐傜傕丄MifM偲儕儃僜乕儉偑丄偙偺嫹嶓晹埵偱愙怗偟偰偄傞偙偲偑帵嵈偝傟丄椉尋媶幒偑撈棫偵峴偭偨夝愅寢壥偑丄偙偺嫹嶓晹埵偺東栿傾儗僗僩偵偍偗傞廳梫傪偦傟偧傟棤晅偗偨偙偲偵側傝傑偡丅
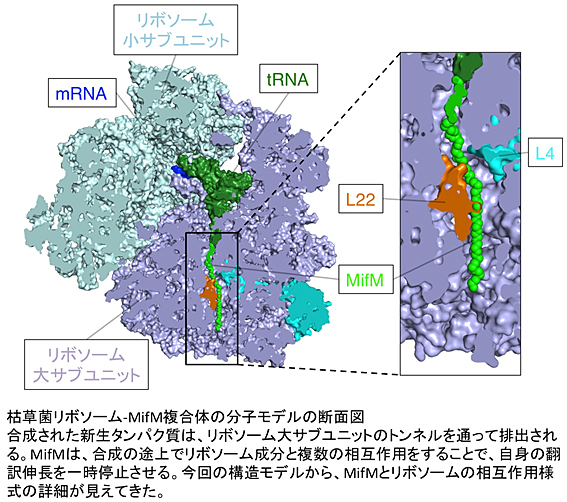
丂師偵巹偨偪偼丄屚憪嬠偲戝挵嬠偺L4偍傛傃L22偺傾儈僲巁攝楍偺堘偄偵拝栚偡傞偙偲偱庬摿堎惈傪寛掕偡傞梫場偲側傝偆傞岓曗偺巆婎傪峣傝崬傒丄偦傟傜偺巆婎傪僞乕僎僢僩偲偟偨曄堎夝愅傪峴偄傑偟偨丅偦偺寢壥丄L22偺90斣栚偺傾儈僲巁巆婎偺庬椶偑屚憪嬠偺儕儃僜乕儉偱婲偙傞東栿傾儗僗僩偺岠棪傪寛掕偡傞忋偱廳梫偱偁傞偙偲傪尒弌偟傑偟偨丅峔憿僨乕僞偐傜偼丄偦偺傾儈僲巁巆婎偑丄嫹嶓晹埵傪宍惉偡傞儕儃僜乕儉偺摿掕偺RNA巆婎傪夘偟偰MifM-儕儃僜乕儉娫憡屳嶌梡偵塭嬁傪梌偊傞壜擻惈偑帵嵈偝傟傑偟偨丅
丂埲慜巹偨偪偼丄MifM偑丄儕儃僜乕儉偺儁僾僠僕儖揮埵峺慺妶惈仸2傪慾奞偟偰偄傞偙偲傪帵偡寢壥傪摼偰偄傑偟偨丅崱夞摼傜傟偨峔憿僨乕僞偼丄巹偨偪偑埲慜採彞偟偨儌僨儖傪棤晅偗傞偲偲傕偵丄偦偺暘巕婡峔偲偟偰丄MifM偑丄儕儃僜乕儉偺妶惈拞怱晅嬤偺摿掕偺RNA巆婎偺攝抲偵塭嬁傪梌偊傞偙偲偱儁僾僠僕儖揮埵斀墳傪慾奞偟偰偄傞偙偲傪帵嵈偡傞傕偺偱偟偨丅
丂崱夞丄峔憿惗暔妛偲堚揱妛傪梈崌偟偨崙嵺嫟摨尋媶偵傛傝丄東栿傾儗僗僩偵昁梫側MifM-儕儃僜乕儉憡屳嶌梡偺徻嵶偑巆婎儗儀儖偱柧傜偐偵側傝丄東栿傾儗僗僩偺暘巕婡峔傪棟夝偡傞廳梫側庤偑偐傝傪摼傞偙偲偑弌棃傑偟偨丅傑偨丄東栿傾儗僗僩偵偍偗傞儕儃僜乕儉偺庬摿堎惈偺栤戣傪棟夝偡傞庤偑偐傝傪摼傞偙偲傕弌棃傑偟偨丅嬤擭丄東栿怢挿斀墳偑懍搙偺娚媫傪敽偭偰恑峴偟丄偦偺懍搙偺娚媫偑丄僞儞僷僋幙偺嬊嵼壔傗惉弉夁掱側偳偵廳梫側栶妱傪扴偭偰偄傞偙偲偑彮偟偢偮柧傜偐偵偝傟偰偒偰偄傑偡丅東栿怢挿偺懍搙挷愡偵娭楢偡傞杮尋媶偼丄彨棃揑偵偼丄僞儞僷僋幙偺抋惗偲惉弉偺栤戣丄偡側傢偪丄堚揱忣曬偐傜惗柦偑惗傒弌偝傟傞偟偔傒偺徻嵶傪棟夝偡傞偙偲傊偲宷偑偭偰偄偔偲婜懸偝傟傑偡丅
仸1 恀惈嵶嬠偼丄偦偺嵶朎昞憌偺峔憿忋偺堘偄偐傜僌儔儉梲惈嬠偲僌儔儉堿惈嬠偵偍偍偒偔暘椶偡傞偙偲偑弌棃傑偡丅偙傟傑偱丄戝挵嬠傗岲擬嬠乮僒乕儅僗僒乕儌僼傿儔僗乯側偳偵戙昞偝傟傞僌儔儉堿惈嬠偺儕儃僜乕儉偺峔憿偼徻嵶偵柧傜偐偵偝傟偰偄傑偟偨偑丄屚憪嬠側偳偺僌儔儉梲惈嬠偺儕儃僜乕儉偺尨巕儗儀儖偱偺峔憿偼柧傜偐偵偝傟偰偄傑偣傫偱偟偨丅
仸2 儁僾僠僕儖揮埵峺慺妶惈丗僞儞僷僋幙偼丄偦偺峔惉惉暘偱偁傞傾儈僲巁摨巑偑丄儁僾僠僪寢崌偲屇偽傟傞寢崌條幃偱嵔忬偵宷偑傟偰弌棃偰偄傑偡丅儕儃僜乕儉偵傛偭偰怗攠偝傟傞偙偺儁僾僠僪寢崌宍惉斀墳偼丄偦偺斀墳條幃偐傜乽儁僾僠僕儖揮埵斀墳乿偲傕屇偽傟丄儕儃僜乕儉偑帩偮偙偺妶惈傪乽儁僾僠僕儖揮埵峺慺妶惈乿偲屇傃傑偡丅
