



生命の始まりを解き明かす――ツメガエル卵母細胞における成熟・受精・その後の運命 生命科学部教授が国際学術誌で高評価 ー Xenopus卵母細胞研究の総説論文公開とExceptional Reviewer選出ー
生命科学部 産業生命科学科・卵細胞発生情報研究室を主宰する佐藤 賢一教授が、国際的学術出版社MDPIが発行する英文学術誌 Cells(1)において、2025年5月に「Exceptional Reviewers List」(2)に選出されました。これは、査読者としての専門性、学術的貢献度、そして迅速かつ高品質な査読活動が高く評価された研究者に与えられるもので、国際学術コミュニティにおける同教授の存在感を示すものです。さらに2025年12月には、同じくMDPIが発行する国際学術誌 Biomolecules(最新インパクトファクター:4.8)(3)において、総説論文 “The Xenopus Oocyte System: Molecular Dynamics of Maturation, Fertilization, and Post-Ovulatory Fate”(ツメガエル卵母細胞系における成熟・受精・排卵後運命の分子動態)が受理・公開されました(4)。本論文は Special Issue: Gametogenesis and Gamete Interaction, 2nd Edition(配偶子形成と配偶子間相互作用の特集号・第2弾)に収載され(5)、筆頭・責任著者として単著で執筆されています。本総説論文では、発生生物学および生殖生物学の分野で長年モデル生物として用いられてきたアフリカツメガエル(Xenopus laevis)の卵母細胞に焦点を当て、減数分裂停止状態から成熟、受精、さらには受精に失敗した場合の細胞死に至るまでの分子・細胞動態を統合的に整理しています。特に、MPF(Cdk1/Cyclin B:卵成熟因子)やMAPK(mitogen-activated protein kinase)カスケード、CPEB(Cytoplasmic polyadenylation element binding protein)依存的翻訳制御といった卵成熟制御機構に加え、哺乳類とは異なる両生類特有の卵活性化シグナル機構(Srcチロシンキナーゼ経路やマトリクスメタロプロテイナーゼ-2活性)について、最新の知見を踏まえて体系的に論じています。また本論文では、未受精卵の運命としてのアポトーシスやネクローシスの分子基盤にも言及し、生殖医療や発生異常研究への応用可能性を示しています。さらに、mRNAマイクロインジェクション、ゲノム編集、in vitro排卵系など、近年進展するXenopus研究技術についても展望を示し、基礎生命科学からトランスレーショナル研究への架橋となる位置づけを明確にしています。佐藤教授は、ツメガエル卵細胞の生物学的機能解明を主軸とした基礎研究に加え、「問いづくり」を重視した教育実践や、生命科学と社会をつなぐコミュニケーションのあり方にも積極的に取り組んでいます。今回の国際的評価および研究成果は、本学生命科学部における研究力と教育力の両立を体現するものとして、今後のさらなる学術的・社会的展開が期待されます。
関連webサイト(URL)
- https://www.mdpi.com/journal/cells
- https://www.mdpi.com/about/announcements/11569
- https://www.mdpi.com/journal/biomolecules
- Biomolecules 2026, 16(1), 22; https://doi.org/10.3390/biom16010022,
- https://www.mdpi.com/journal/biomolecules/special_issues/G9NC3XYLG1
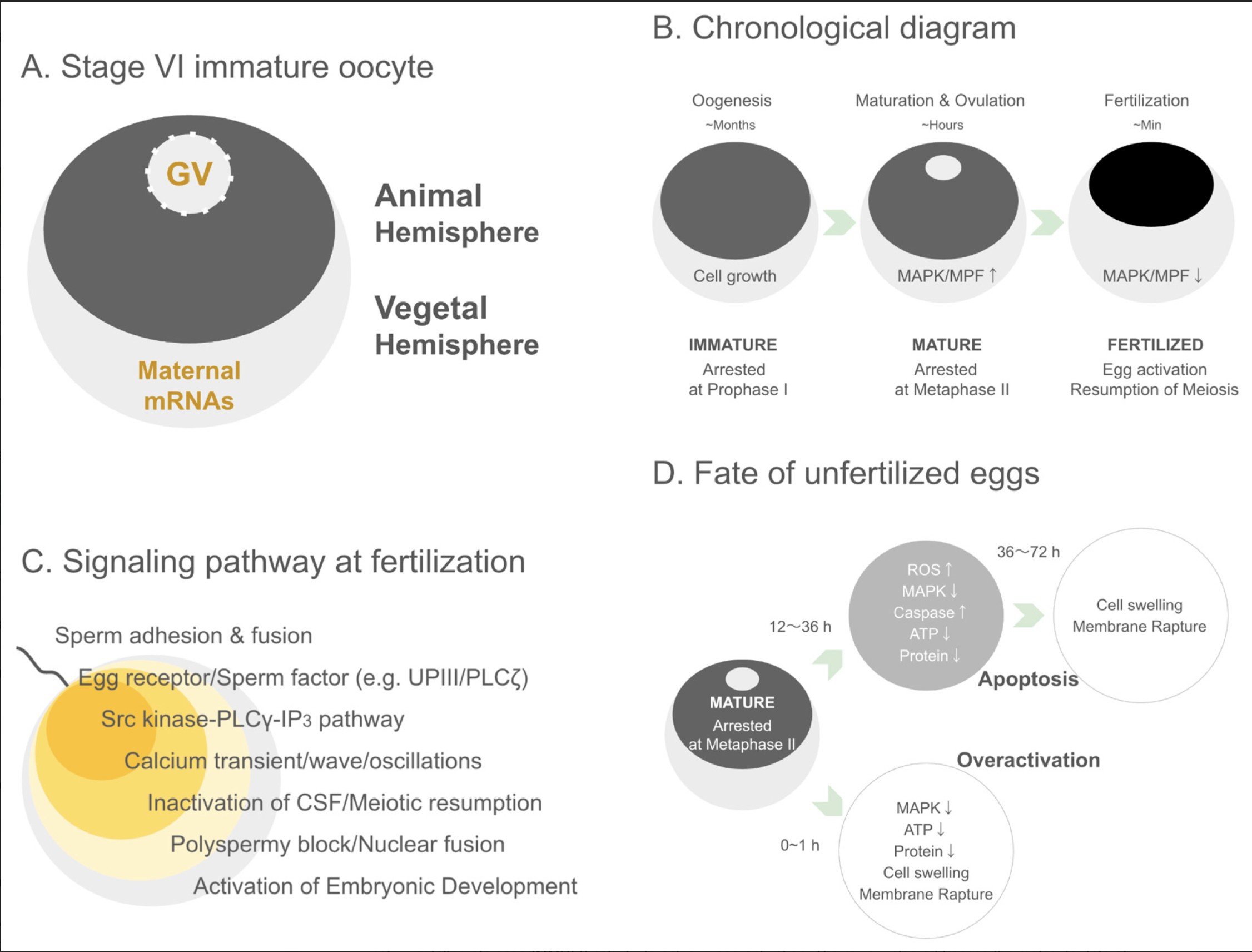
ツメガエル卵母細胞・卵における構造、発生過程、シグナル伝達、および排卵後の運命
本図は、ツメガエル(Xenopus)の卵母細胞および卵における構造的特徴、発生段階の推移、受精時のシグナル伝達機構、ならびに受精に至らなかった場合の細胞運命を模式的に示したものです。
(A)ステージVI未成熟卵母細胞 完全に成長したステージVI卵母細胞は、動物半球と植物半球からなる明瞭な極性構造を有しています。色素の濃い動物半球には生殖胞(germinal vesicle:GV)が存在し、一方、植物半球には受精後の初期胚発生の方向性を制御する母性mRNAが豊富に蓄積されています。この極性は、胚の体軸形成において重要な役割を果たします。
(B)時間軸に沿った発生過程 卵母細胞は、数か月に及ぶ卵形成期において成長を続けながら、減数分裂第I期前期で停止しています。その後、ホルモン刺激を受けることで、数時間のうちに減数分裂成熟と排卵が進行し、MAPKやMPFといった細胞周期制御因子の活性が上昇し、減数分裂第II期中期で再び停止します。受精が起こると、これらの活性は急速に低下し、卵は活性化されて減数分裂を再開し、胚発生が始まります。
(C)受精時のシグナル伝達経路 受精時には、精子の付着・融合をきっかけに、卵表面の受容体や精子由来因子(ウロプラキンIIIやPLCζなど)が作動し、細胞内シグナル伝達が開始されます。特に、Srcキナーゼ–PLCγ–IP3経路の活性化により、特徴的なカルシウム濃度変動(波状・振動的応答)が誘導されます。これにより、CSF(細胞質成熟因子)が不活化され、減数分裂が再開します。カルシウム依存的な反応は、多精子受精の防止、前核融合、そして胚発生の開始を促進します。
(D)未受精卵の運命 排卵後に受精が起こらなかった卵は、時間の経過とともに劣化していきます。12〜36時間後には、MAPK活性やATP量、タンパク質量の低下とともに、活性酸素(ROS)の増加やカスパーゼ活性化が進行し、最終的にアポトーシスへと至ります。その結果、36〜72時間後には細胞膨張と細胞膜破綻が起こります。一方で、排卵後0〜1時間以内に、過剰活性化(オーバーアクティベーション)が生じる場合もあり、この場合はMAPK活性の急激な低下、ATP枯渇、細胞質の急速な劣化、早期の膜破綻が引き起こされます。これらの異なる経路は、排卵後未受精卵が持つ代謝的・シグナル伝達上の脆弱性を示しています。